1. ヒト造血幹細胞ゲノム編集によりムコ多糖症 (MPS) I型の表現型修復を実現
[出典] Human genome-edited hematopoietic stem cells phenotypically correct Mucopolysaccharidosis type I. Gomez-Ospina N [..] Porteus MH. bioRxiv. 2018 Sep 4.
- リソソーム内の酵素の不全による広範な遺伝病には効果的療法が無い。Stanford University School of Medicineを始めとする米国・デンマークの研究グループは今回、セーフハーバーを利用したゲノム編集療法の可能性を示した。
- ヒトCD34陽性造血幹細胞・前駆細胞 (HSPCs)において、ヒトゲノムのセーフハーバーとされるCCR5 遺伝子座に、リソソーム酵素のイズロニダーゼ (iduronidase, IDUA)をCRISPR/Cas9を利用して挿入し、IDUAを過剰発現させた。
- ゲノム編集したHSPCsは自己複製能と多分化能を長期間維持し、酵素を高発現し、免疫不全MPS Iモデルマウスの生化学的特性および表現形質を正常化した。
2. 転写共役型相同組換え修復(Transcription-Associated Homologous Recombination Repair; TA-HRR)の分子機構解明
[出典] Human Rad52 Promotes XPG-Mediated R-loop Processing to Initiate Transcription-Associated Homologous Recombination Repair. Yasuhara T, Kato R, Hagiwara Y, Shiotani B, Yamauchi M, Nakada S, Shibata A, Miyagawa K. Cell. 2018 Sep 18.
- 東大、群大、がん研 (NCC)、長崎大、阪大の研究グループは今回、放射線損傷や抗癌剤などにより転写活性化領域に誘導されるDNA損傷、DSB (二本鎖DNA切断)、の~5%に発生するTA-HRRの分子機序を、CRISPR-Cas9によりRad52を欠損させた網膜色素上皮細胞(RPE細胞)での実験などにより、明らかにした。
- 転写活性化領域におけるDSBは、DNA-RNAハイブリッドからなるR-ループを形成し、Rad52、続いて、 CtIP、BRCA1およびエンドヌクレアーゼXPGをリクルートする。DSB領域に集積したBRCA1によるRIF1-53BP1複合体の阻害と、XPGによるR-ループ除去を経て、DSB 末端の削り込み (end resection)が進行し、DNA修復因子RPA(Replication Protein A)とRad51がリクルートされる。また、ATMを介したCtIPとBRCA1のリン酸化が進行する。その結果、TA-HRRが進行するに至る。
- 電離放射線に起因するDSBの70%以上がNHEJによって修復されるがゲノム異常を生じることは稀である。しかし、DSB修復の~5%を担うTA-HRRがNHEJにスイッチすると、顕著な染色体異常が発生する。研究グループは、Rad52欠損などTA-HRR経路の阻害が染色体異常を亢進することも確認した。
3. ブタ内在レトロウイルス (PERVs):細胞株、各品種および器官におけるコピー数定量化
[出典] Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Fiebig U, Fischer K, Bähr A, Runge C, Schnieke A, Wolf E, Denner J. Xenotransplantation. 2018 AUG 18.
- PERVsはブタゲノムに内在しヒトへのブタ臓器異種移植における障害となっている。Robert Koch Instituteなどドイツの研究チームは、ブタ細胞株 (PK15とPTK75)と個体 (アーヘン/ゲッチンゲン/ブラック・ミニブタ;野生および遺伝子操作 German landraceブタ)におけるPERVsをddPCRで定量し、コピー数が46-117の範囲で変動し、また、同一個体ないでも器官に依存して変動していることを見出した。これらは、CRISPR/Cas9によるPERVプロウイルス不活性化処理に有用な情報である。
4. [レビュー] 癌のin vivo機能ゲノミクスの定量化と多重化に向けて
[出典] REVIEW: Towards quantitative and multiplexed in vivo functional cancer genomics. Winters IP, Murray CW, Winslow MM. Nat Rev Genet. 2018 Sep 28.
- ヒト癌の大規模なシーケンシングプロジェクトから得られるゲノム変異の機能をin vivoで解析することが可能になってきた。特に、CRISPR-Cas9技術により、複雑な体細胞変異モデルを迅速に作出可能になったことが貢献している。
5. CRISPR gRNAリボスイッチの提案
[出典] Engineering CRISPR guide RNA riboswitches for in vivo applications. R Galizi, A Jaramillo A. Curr Opin Biotechnol. 2018 Sep 25.
- 低分子や光などで活性を誘導可能なCRISPR-Casシステムが開発されてきたが著者らは今回、リボスイッチの技術を概観した上で、リボスイッチ技術とCRISPR-Cas技術の融合を提案した。すなわち、gRNAにリボスイッチとして機能するRNAを結合し、このリボスイッチを低分子やトリガーとなるRNAを介してオンオフすることで、gRNAの標的への結合ひいてはCRISPR-Casの編集活性を誘導・調節する。
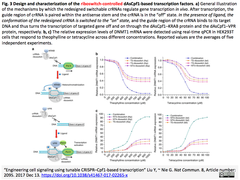
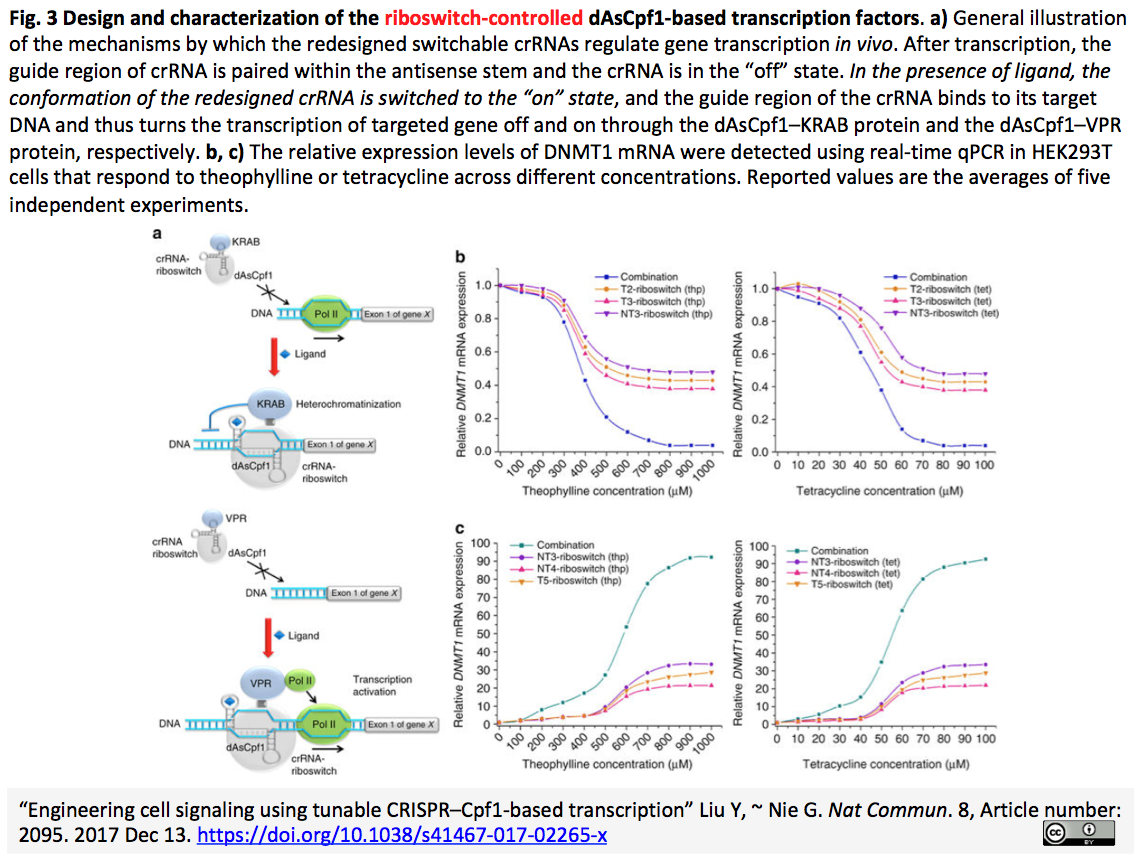
- 関連crisp_bio記事:2017-12-15 CRISPR-Cpf1をベースにした調節可能な転写因子により細胞内シグナル伝達を制御する - crRNA-リボスイッチによる転写抑制と活性化の誘導 (Fig.3を以下に再掲)
6. CRISPR-Cas9遺伝子編集はエクソン・スキッピングを誘導する
[出典]CRISPR/Cas9-mediated genome editing induces exon skipping by complete or stochastic altering splicing in the migratory locust. D Chen, JX Tang, B Li, L Hou, X Wang, L Kang - BMC Biotechnology. 2018 Sep 25.
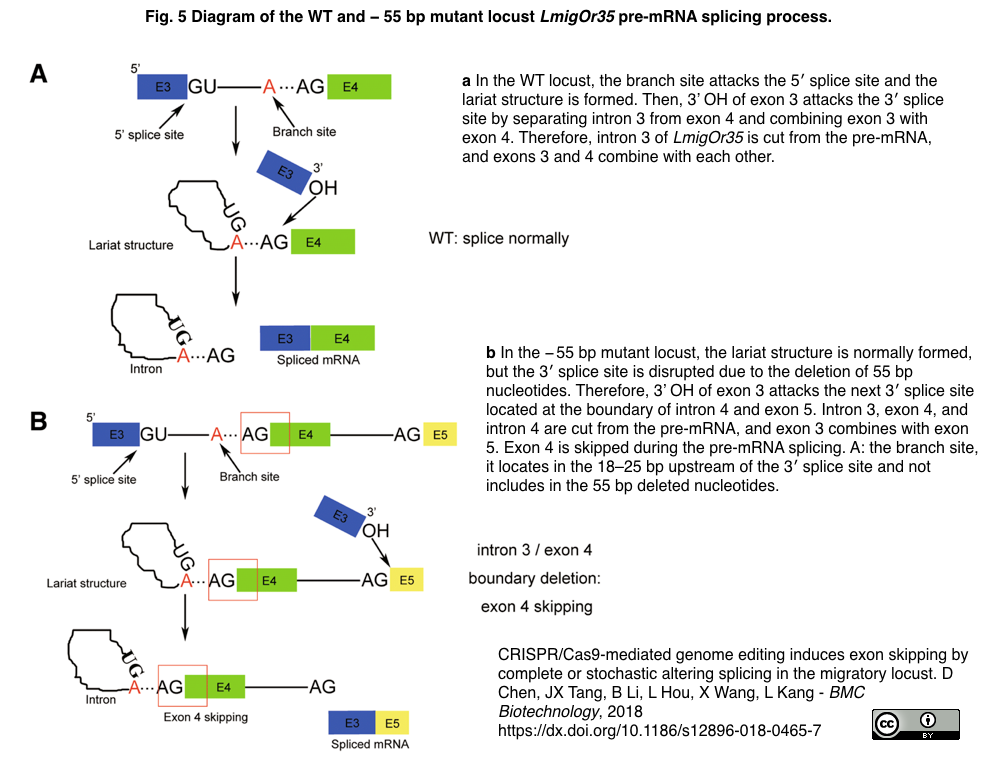
- 長いイントロンの独特なスプライシング機構を帯びたゲノムサイズ6.5Gbのトノサマバッタ (Locusta migratoria)での実験; 嗅覚受容体遺伝子LmigOr35の第3イントロンと第4エクソンの境界における55 bp欠損 (イントロン 15 bp; エクソン 40 bp)に伴う3'スプライス部位破壊により第4エクソンが100%スキップされる一方で、第4エクソンにおける22 bpの欠損は確率的な第4エクソン・スキッピング (mRNAの~26%)を誘導した (100%スキップをもたらすスプライシング機構:原論文Fig. 5引用下図参照)。
- CRISPRメモ_2018/09/04- 3. DMD患者由来iPS細胞から誘導した筋芽細胞において、CRISPR-Cas9エクソンスキッピングにより、ジストロフィンタンパク質発現を修復
- CRISPRメモ_2018/08/19 - 1. CRISPR-SKIP:一塩基エディタによるプログラム可能な遺伝子スプライシング
- "CRISPR/Cas9-mediated genome editing induces exon skipping by alternative splicing or exon deletion" Mou H, Smith JL [..] Moore MJ, Weng Z, Xue W. Genome Biol. 2017 Jun 14
- "Random splicing of several exons caused by a Single Base change in the target exon of CRISPR/Cas9 mediated gene knockout" Kapahnke M, Banning A, Tikkanen R. Cells. 2016 Dec 14.
7. TCPファミリー転写因子に属するBRANCHED1とBRANCHED2のポプラ・ホモログの機能
[出典] CRISPR/Cas9-mediated knockout of Populus BRANCHED1 and BRANCHED2 orthologs reveals a major function in bud outgrowth control. Muhr M, Paulat M, Awwanah M, Brinkkötter M, Teichmann T. Tree Physiol. 2018 Sep 28.
- CRISPR/Cas9遺伝子機能喪失実験により、ポプラにおいて、BRANCHED1はシロイヌナズナと同等の機能を担うが、BRANCHED2はシロイヌナズナと異なり、芽の伸長の調節に決定的な因子であることを同定。

コメント