[注] 本稿はCRISP_SCIENCEのCRISPR関連ツイート(2017年12月3日)に準拠しています。
1.CRISPR/Cas9タンパク質とドナーDNAを接合子にエレクトロポレーションすることで遺伝子編集ラットを作出
- [出典] "Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation" Remy S, ~ Anegon I. Sci Rep. 2017 Nov 29;7(1):16554
- マイクロインジェクションにかわりエレクトロポレーションにより、NHEJを介した15−50%のノックアウト効率、〜100-nt ssODNをドナー・テンプレートとして利用することで、25−100%のノックイン効率を実現。より長いssODNまたはss/dsDNAの挿入は実現せず。
2.効率的なゲノム編集のためにブタ卵母細胞にCRISPR/Cas9 mRNAをマイクロインジェクションするタイミングは、電気刺激による活性化の6時間後(前核形成時)が最適
- [出典] "Timing of CRISPR/Cas9-related mRNA microinjection after activation as an important factor affecting genome editing efficiency in porcine oocytes" Sato M, ~ Miyoshi K. Theriogenology. Available online 26 November 2017
- 先行研究では電気刺激活性化直後にマイクロインジェクションすることでゲノム編集を実現したが、モザイク状の胚盤胞が発生し、また、編集効率は50%に止まった。今回、編集効率は69%に達したが、モザイク現象は減少しなかった。
3.BE3によるアフリカツメガエル胚の一塩基編集
- [出典] "Targeted Base Editing via RNA-Guided Cytidine Deaminases in Xenopus laevis Embryos" Park DS, Yoon M, Kweon J, Jang AH, Kim Y, Choi SC. Mol Cells. 2017 Nov 30;40(11):823-827.
- チロシナーゼを標的とするBE3 RNPにより一塩基変換を20.5%の効率で達成し、オフターゲット作用無しに皮膚白皮症の表現型を生成;tp53遺伝子を標的とする一塩基変換も確認
4.CRISPRイメージングにより植物細胞内テロメア動態を観察
- [出典] "Live-cell CRISPR imaging in plants reveals dynamic telomere movements" Dreissig S, ~ Puchta H, Houben A. Plant J. 2017 Aug;91(4):565-573. Published online 2017 Jul 14.
- Nicotiana benthamianaの生細胞においてテロメアの反復配列を標的とするsgRNAと、Sp-dCas9またはSa-dCas9にeGFPまたはmRuby2を融合したコンストラトにより、テロメアの動態をin situ観察(Figure 1とFigure 4参照)。また、テロメア反復配列結合タンパク質TRB1の蛍光標識を組み合わせて、DNAータンパク質間相互作用観察も観察(Figure 5)
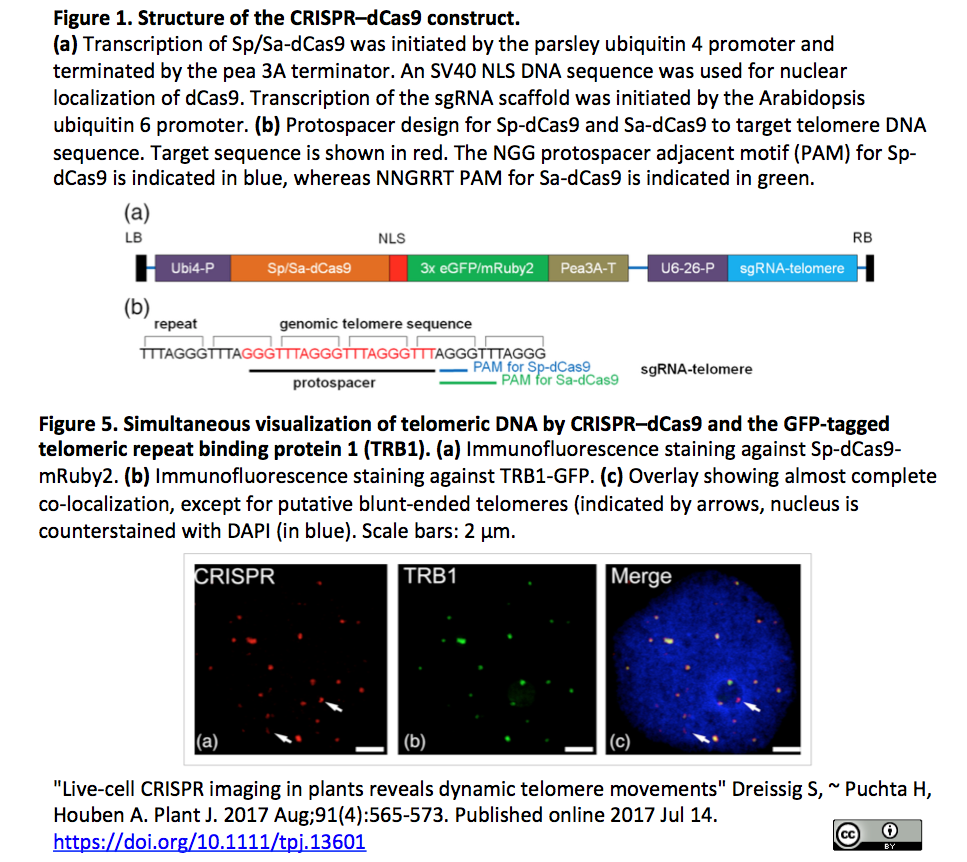
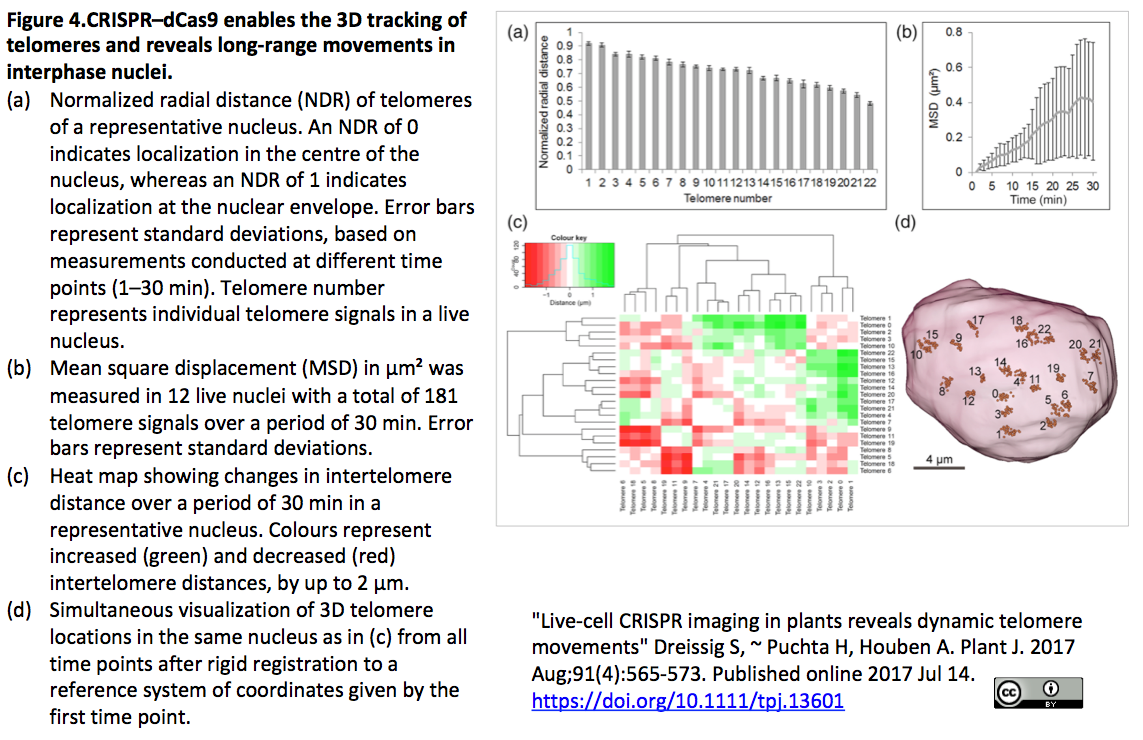
- [参考:特定の染色体を丸ごと可視化] "Painting a specific chromosome with CRISPR/Cas9 for live-cell imaging" Zhou Y, Wang P, Tian F, Gao G, Huang L, Wei W, Xie XS. Cell Res. 2017 Feb;27(2):298-301. Published online 13 January 2017
5.Arabidopsis thalianaにおいてCRISPR/Cas9が誘導するDSBの修復モードは内在配列に依存する
- [出典] "Endogenous sequence patterns predispose the repair modes of CRISPR/Cas9-induced DNA double-stranded breaks in Arabidopsis thaliana" Vu GTH, Cao HX, Fauser F, Reiss B, Puchta H, Schubert I. Plant J. 2017 Oct;92(1):57-67. First published 14 August 2017.
- CRISPR/Cas9がA. thalianaの三種類の遺伝子座に誘導する変異スペクトルをロング・アンプリコン・ライブラリーのディープ・シーケンシングで解析し、1-bp〜<1000-bpの範囲そしてまたは極めて短い挿入、NHEJによる1-kbpを超える欠失(NHEJに依る)および5-bpから100-bpを超える挿入を伴う欠失(synthesis-dependent strand annealing (SDSA)様の機構に依る)が、もっとも高頻度で発生し、また、変異スペクトルがDSBサイト周囲のDNA配列(反復配列、cis 相同配列、マイクロホモロジーなどの存否・配置)に依存することを見出した。
6.腸炎菌(Salmonella enterica serovar Enteritidis)の遺伝解析とCRISPRタイピングによって、腸炎菌の家禽と豚からヒトへの伝播の可能性が示唆された
- [出典] "Genetic analysis and CRISPR typing of Salmonella enterica serovar Enteritidis from different sources revealed potential transmission from poultry and pig to human" Li Q, ~ Jiao X. Int J Food Microbiol. Available online 28 November 2017.
- 腸炎菌329株を解析し18種類のCRISPRタイプ(Enteritidis CRISPR types, ECTs)を同定
7.アンチセンスRNAによるcas遺伝子のサイレンシングに拠り、バクテリアファージ耐性Streptococcus thermophilusを作出
- [出典] "Generation of bacteriophage-insensitive mutants of Streptococcus thermophilus using an antisense RNA CRISPR-Cas silencing approach" McDonnell B, Mahony J, Hanemaaijer L, Kouwen TRHM, van Sinderen D. Appl Environ Microbiol. 2017 Nov 27.
- スタータとして食品産業に利用されるS. thermophilusへのバクテリオファージ感染の防止;CRISPR-mediated Bacteriophage Insensitive Mutants (BIMs)に対してバクテリオファージが容易に耐性を獲得することから、CRISPRシステムに依存しない手法を開発

コメント