1. SpCas9活性の深層学習モデルDeepCas9 (2019-09-20, -11-10, -11-26 追記)
[出典] "SpCas9 activity prediction by DeepCas9, a deep learning-based model with unparalleled generalization performance" Kim HK, Kim Y [..] Kim HH. bioRxiv 2019-05-15. > "SpCas9 activity prediction by DeepSpCas9, a deep learning–based model with high generalization performanc" Sci Adv. 2019-11-06
- 延世大学校医科大学の研究グループは、先行研究にてAsCpf1活性の深層学習モデルDeepCpf1を構築した手法 (crisp_bio *1)に準拠し、SpCas9活性の深層学習モデルDeepCas9を構築・公開した。
- 15,656組のsgRNAsと標的配列 (合成DNA)を導入したHEK293T細胞ライブラリを構築し、続いて、SpCas9を導入し、SpCas9の活性 (indels変異頻度)を測定し、12,832組のデータに基づいてニューラルネットワーク(Convolutional Neural Network, CNN)に基づく深層学習モデルを構築し、542組のSpCas9活性データで検証した。
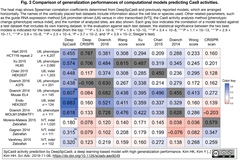
- また、SpCas9活性の10種類の公開データセットを利用して、DeepCRISPR (crisp_bio *2)を含む既存の予測モデル8種類と性能を比較し、DeepCas9が最も高精度とした (in Delphi crisp_bio *3;Linモデル *; FORECasT *5)は比較対象に含まれていない) [Sci Adv.論文掲載Fig. 3引用下図参照 (bioRxiv.投稿掲載Fig.3と同一)]
- DeepCas9はhttp://deepcrispr.info/DeepCas9にて公開
参考crisp_bio記事とツイート
- (*1) CRISPRメモ_2018/02/04 [第3項目] CRISPR-Cpf1 gRNA活性予測精度を、深層学習 (deep learning) により向上
- (*2) CRISPRメモ_2018/07/02-1 [第7項目] DeepCRISPR:深層学習によるCRISPR gRNAの最適設計
- (*3) 2018-11-08 機械学習モデルにより一塩基編集技術を磨く - ディープラーニングに基づくモデルにより、NHEJ/MMEJを介した1-bp挿入と病因変異修復を実現 (inDelphi)
- (*4) CRISPRメモ_2018/09/16 [第9項目] ディープラーニングによるCRISPR-Cas9遺伝子編集のオフターゲット予測
- (*5) CRISPRメモ_2018/08/27 [第1項目] Cas9が誘導するDSBの修復過程で生成される変異は、周囲の配列から予測可能
- 2019/09/20 DeepHF: SpCas9とその高精度版2種類に最適なgRNAsを設計する深層学習モデルを構築・公開
- (参考) CRISPRメモ_2018/08/11 [第1項目] CRISPRゲノム編集の精度は標的サイトによって大きく変動する
- Phys.orgのツイート
 CRISP_SCIENCE@ScienceCrisp
CRISP_SCIENCE@ScienceCrispA deep learning-based model DeepSpCas9 to predict SpCas9 activity https://t.co/yS3ocIRaWY via @physorg_com
2019/11/26 09:17:05
2. [レビュー] CRISPR Cpf1 (Cas12a)タンパク質の構造、機能およびゲノム編集への応用
[出典] "CRISPR Cpf1 proteins: structure, function and implications for genome editing" Safari F [..] Ghasemi Y. Cell Biosci 2019-05-09.
- Shiraz University of Medical Sciences (イラン)とCase Western Reserve University (米国)のグループによる引用文献156編21ページにわたるレビュー
3. ウサギにおいてBE3による塩基Gの直下に位置するC (GCコンテクスト)のTへの変換効率を向上
[出典] "Improved base editor for efficient editing in GC contexts in rabbits with an optimized AID-Cas9 fusion" Liu Z [..] Lai L, Li Z. FASEB J 2019-05-09.
- 吉林大学の研究グループは、BE3のrAPOBEC1を活性化誘導シチジンデアミナーゼ (AID)に変更することで、ウサギにおいて11-ntウインドウ幅におけるC-to-T変換効率が向上することを示した。
- 特に、enhanced AID (eAID)とBE4maxの組み合わせ[eAID-BE4max*]により、Tyrp.R299H変異におけるGCコンテクストのC-to-T変換効率がBE3による16.67%から83.33%に向上することを示した。さらに、eAIDの変異体eAID (N51G)とBE4maxの組み合わせ[eAID (N51G)-BE4max] によりバイスタンダー変異が抑制されることを示した。
- eAIDは,C末端の核輸出シグナル(P128X9)を欠失させ、同時に3つの高活性化変異 (K10E, T82I, E156G)を導入したAIDである。
- (*) CRISPRメモ_2018/05/30-3 [第1項目] 塩基エディターBE4とABEを、BE4max / AncBE4max / ABEmaxへと強化
4. S. epidermidisにおけるCas10-Csm複合体によるサイクリックオリゴアデニル酸合成の調節機構
[出典] "Regulation of cyclic oligoadenylate synthesis by the S. epidermidis Cas10-Csm complex" Nasef M [..] Hatoum-Aslan A, Dunkle JA. RNA 2019-05-10.
- タイプⅢ CRISPR-Casシステムは、サイクリックオリゴアデニル酸(cOA)を合成した非特異的なRNA分解機能も備えていることが知られており、いくつかのバクテリアとアーケアで分子機構が解析されている*。
- University of Alabamaの研究グループは今回、S. epidermidisのタイプIII-A CRISPR-CasにおけるcOA合成・調節の機構を解析した:3-6 ntのcOAはMgイオン依存で合成される;cOA合成の活性化は、crRNAと標的RNAとの間の1-nt ミスマッチで阻害される;cOA合成は、Csm3を介した標的RNAの切断と、拮抗する
- (*) 参考crisp_bio記事:2019-03-24タイプIII CRISPR-Casの非特異的RNA分解活性を仲介するセカンドメッセンジャーを分解する'ring nuclease'
5. LinTIMaT: CRISPR-Cas9バーコード編集とscRNA-seqの融合による細胞系譜追跡
[出典] "Single-cell Lineage Tracing by Integrating CRISPR-Cas9 Mutations with Transcriptomic Data" Zafar H, Lin C, Bar-Joseph Z. bioRxiv 2019-05-07.
[改訂版] Nat Commun2020-06-16. [2020-07-04 LinTIMaT: バーコード上のCas9誘導変異データとscRNA-seqデータを統合した細胞系譜再構成法]
[改訂版] Nat Commun2020-06-16. [2020-07-04 LinTIMaT: バーコード上のCas9誘導変異データとscRNA-seqデータを統合した細胞系譜再構成法]
- Carnegie Mellon Universityの研究チームは、DNA断片にCRISPR/Cas9がランダムに誘導するindels変異のパターンの変遷データに基づく細胞系譜の再構成と、scRNA-seqによる細胞型の同定とを組み合わせて細胞運命マップを描き出すscGESTALT (*)をはじめとする手法について、ランダム変異データの処理の仕方に課題があるとし、最尤法の枠組みで変異データとトランスクリプトームデータを統合していくことで細胞系譜を描き出すLinTIMaT (Lineage Tracing by Integrating Mutation and Transcriptomic data)法を開発し、scGSETALTの実験で利用されたゼブラフィッシュの2種類のデータセットにて、その性能を検証した。
- (*) 2018-04-12細胞バーコーディングとscRNA-seqを一体化して、細胞型の同定と細胞系譜再構築の一石二鳥を実現 [第3項目]

コメント