[出典] "Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity" Richter MF, Zhao KT [..] Doudna JA, Liu DR. Nat Biotechnol 2020-03-16
[ABE関連資料] crisp_bio 2020−03−18 crisp_bioコレクション: ABE
[関連crisp_bio記事] crisp_bio 2020-03-18 Beam Therapeutics、ABE7.10をABE8へと進化 [注: 異なる指向性進化法による異なる観点からのABE7.10のバージョンアップ]
[ABE関連資料] crisp_bio 2020−03−18 crisp_bioコレクション: ABE
[関連crisp_bio記事] crisp_bio 2020-03-18 Beam Therapeutics、ABE7.10をABE8へと進化 [注: 異なる指向性進化法による異なる観点からのABE7.10のバージョンアップ]
背景
- DR Liuらは先行研究で、不活性化したCas9 (dCas)にE. coli由来アデノシン脱アミノ化酵素 (TadA)から導出したTadA変異型 (TadA*)を融合したABE7.10は、A•T-to-G•C変換を実現した。
- また、ABEの標的可能範囲を広げるために、SpCas9以外のCasエフェクタの利用を試みたが、編集効率は極めて低いか良くても中程度に留まった。
- その原因を、アデニンの脱アミノ化が低速であることと、エフェクターによってはDNAへの結合時間が短いことによると想定して、実験を進めた。
成果
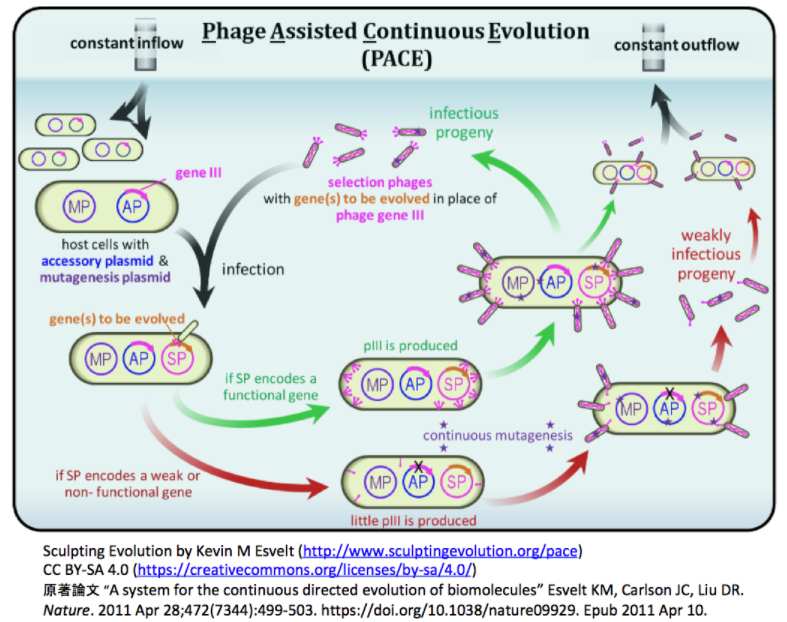
- Broad Institute/Harvard UとUC Berkeleyなどの研究グループは、先行研究で開発しCBEの性能向上に利用した連続的な指向性進化法PACE (Phage-Assisted Continuous Evolution) [下図参照]とその派生型 PANCE (Phage-Assisted Non-Continuous Evolution) [参考文献参照]を利用して、ABE7.10のTadA*を指向性進化させた。
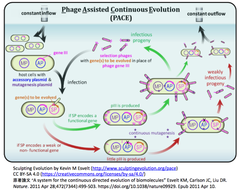
- その結果、脱アミノ化反応速度がTadA*の590倍のTadA-8eを同定し、また、TadA-8eがSpCas9, SaCas9, LbCas12a, enAsCas12a, SpCas9-NG, SaCas9-KKH, CP1028-SpCas9およびCP1041-SpCas9のいずれと組み合わせても、塩基エディティングの活性を示することを見出し、ABE8eに至った。
- SpCas9以外のエフェクターと組み合わせ可能になったことから、A•T-to-G•C変換を実現可能な標的可能な領域が拡大された。
- 一方で、ABE8eはオフターゲットのDNA編集とRNA編集を亢進することになった。この問題は、先行研究でオフターゲット編集抑制効果があるとされた一連の変異を評価し、TadA-8eドメインにV106Wを導入することで、ABE8eのオンターゲット活性を損なうことなく、ABE7.10よりさらに抑制することで、解決した。
- さらに、HEK293T細胞において、ABE8eによって胎児型ヘモグロビンの発現を亢進する天然変異のBCL11AのエンハンサーまたはHBGプロモーターへの導入を、効率54%で実現した。これらはABE7.10では極めて編集効率が低い標的であった。
PANCE 参考文献
- "Phage-Assisted Evolution of Bacillus methanolicus Methanol Dehydrogenase 2" Roth TB, Woolston BM, Stephanopoulos G, Liu DR. ACS Synth Biol 2019-03-11.
- PACE法関連: crisp_bio 2019-07-24 一塩基編集 (CBE)、ファージに依存する持続的進化法 (PACE)により、進化続く
- PANCE法関連: crisp_bio 2020-02-13 Liuグループ、グアニン (G)を含まない配列をPAMとするSpCas9変異体をファージによる指向性進化法で実現

コメント