1.[特許]遺伝子産物の発現調節を目的とするCRISPR-Casシステムと手法、構造情報および誘導可能なモジュラーCasエンドヌクレアーゼ
- "CRISPR-CAS SYSTEMS AND METHODS FOR ALTERING EXPRESSION OF GENE PRODUCTS, STRUCTURAL INFORMATION AND INDUCIBLE MODULAR CAS ENZYMES" US2018/0066242 A1 PubDate 03/08/2018
- Inventors: Zhang F, Zetsche B
- Assignee: Broad, MIT
2.CRISPR SAM gRNA発現アレイを迅速生成する新奇ベクターシステムを開発
- "Simple method for assembly of CRISPR synergistic activation mediator gRNA expression array" Vad-Nielsen J, Nielsen AL, Luo Y (Aarhus U). J Biotechnol. Volume 274, 20 May 2018, Pages 54–57.
- ヒト乳癌細胞株において、4種類の転写因子 (TWIST, SNAIL, SLUGならびにZEB1)の同時活性化を実現
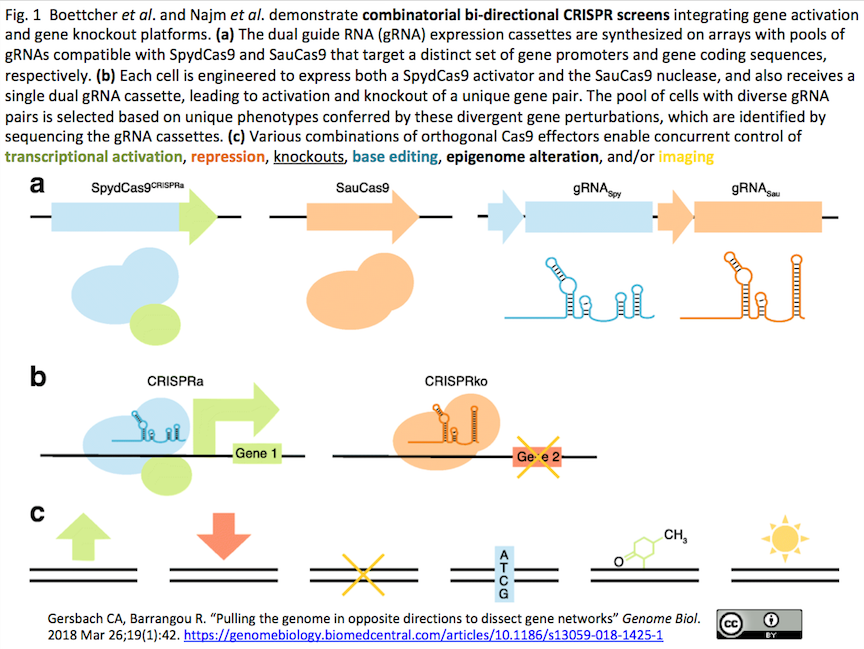
3.[リサーチハイライト]CRISPRa, CRISPRi, CRISPR KO, BE(Base Editor), エピゲノム改変、可視化などを複合することで複雑な遺伝子ネットワークを解読していく時代
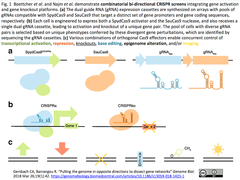
- "Pulling the genome in opposite directions to dissect gene networks" Gersbach CA (Duke U), Barrangou R (North Carolina State U). Genome Biol. 2018 Mar 26;19(1):42.
- 互いに直交するCRISPR-Casシステムを組み合わせたスクリーンに取り組んだ2編の研究論文(UCSFのBoettcher等;BroadのNajm等)をハイライト(下図:不活性化したSpyCas9 (SpydCas9)によるCRISPRaとSauCas9によりCRISPR KOの組み合わせからは始まる多様な遺伝子編集の組み合わせ例)
- Boettcherr論文のcrisp_bio該当記事:2018/01/16-3.遺伝子間相互作用の方向依存性を、遺伝子活性化と遺伝子ノックアウトを組み合わせることで明らかに
- Najm論文のcrisp_bio該当記事:2017/12/24-2.SaCas9とSpCas9の併用による効率的なコンビナトリアル遺伝子スクリーンを実現
4.CRISPR-Cas9システムによりEscherichia coli BL21ゲノムに点変異を生成
- "Making point mutations in Escherichia coli BL21 genome using the CRISPR-Cas9 System" Wang X, He J, Le K (復旦大学). FEMS Microbiol Lett. 27 March 2018.
- CRISPR-Cas9によるdsDNA切断からのHDRを介した点変異導入は、標的サイトがプロとスペーサーかPAM領域にない限り、Cas9により再び切断されるリスクを伴う。復旦大学の研究チームは今回、2段階のCRISPR-Cas9編集法を開発して、proB遺伝子における1塩基置換c.319A > Gを実現した。この点変異導入により、E. coliのプロリン過剰生産が実現し浸透圧ストレスに対する耐性が亢進した。
- 2段階CRISPR-Cas9編集法:(1) CRISPR/Cas9により、長さ20 bpの内在プロトスペーサーを同じ長さの合成プロトスペーサーで置換; (2) 合成プロトスペーサーを認識するsgRNAと、319A > G変異を帯びた内在プロトスペーサー・ホモログをノックイン
5.最適化Target-AIDシステムによるゼブラフィッシュの効率的1塩基置換を実現
- "Optimized Target-AID system efficiently induces single base changes in zebrafish" Lu X, ~ Qin W, Lin S. J (UCLA, 北京大学) Genet Genomics. Available online 26 March 2018.
- Target-AID関連crisp_bio関連記事:2017/04/27-1 [論文] Target-AID: 二本鎖DNA(dsDNA)切断とドナーDNA(テンプレート)を必要としない点変異導入法
6.[レビュー]癌生物学におけるトランスポゾンによる機能ゲノミクスとCRISPR/Cas9による進展
- "Advances in functional genetic screening with transposons and CRISPR/Cas9 to illuminate cancer biology" O’Donnell KA (UT Southwestern Medical Center). Curr Opin Genet Dev. Volume 49, April 2018, Pages 85–94.
- Sleeping Beauty (SB)とPiggyBac (PB);PiggyBac突然変異誘発スクリーン、ex vivoスクリーン、CRISPR/Cas9突然変異誘発ツールボックスとの組み合わせ
7.遺伝子治療:CRISPR/Cas9システムをアデノウイルスで送達することで、ヒトα1-アンチトリプシン (hAAT) cDNAをin vivoノックイン
- "Targeted in vivo knock-in of human alpha-1-antitrypsin cDNA using adenoviral delivery of CRISPR/Cas9" Stephens CJ, Kashentseva E, Everett W, Kaliberova L, Curiel DT (Washington U, Saint Louis). Gene Ther. 2018 Mar 27.
- AATの欠損はα1-アンチトリプシン欠損症を、異常なAATの肝臓への蓄積は肝疾患を、引き起こす。WUSTLの研究チームは今回、遺伝子組み換えアデノウイルスAd5を送達手段としてマウス肝臓のROSA26セーフ・ハーバー領域にhAAT遺伝子をノックインすることで、マウス血清中のhAATを長期間 (200日以上)高いレベルに維持。
8.シロイヌナズナで多様な機能を担うERECTRA遺伝子ファミリー (ERfs)のイネにおける機能を、進化系統解析とCRISPR/Cas9機能喪失実験から推定
- "Phylogenetic and CRISPR/Cas9 studies in deciphering the evolutionary trajectory and phenotypic impacts of rice ERECTA genes" Zhang Y, Li S, Xue S, Yang S, Huang J, Wang L (南京大学). Front Plant Sci. Accepted 26 Mar 2018.
- 進化系統解析からERfsのイネを含む種間保存性が高いことを見出し、CRISPR/Cas9実験からイネのER1とER2遺伝子の欠損による形態変化を同定した。
9.[レビュー]CRISPR/Cas革命RNAワールドに至る:植物学者の新たなスイス・アーミーナイフ、Cas13
- "The CRISPR/Cas revolution reaches the RNA world: Cas13, a new Swiss Army knife for plant biologists" Wolter F, Puchta H (Botanical Institute, Karlsruhe Institute of Technology). Plant J. 2018 Mar 25.
- Cas9とCas12(Cpf1)によりDNA工学に革命が起きたように、Cas13a(C2c2)とCas13bによりRNA工学に革命が起きた。ssRNAの切断に始まり、植物における遺伝子発現の転写後調節、ウイルス感染防御、RNAのin vivo可視化、RNA1塩基置換が報告されている。また、DNAの編集とRNAの編集を組み合わせた植物代謝工学も始まっている。
- [注]Cas13関連crisp_bio記事表示:URL欄に"Cas13 site:http://crisp-bio.blog.jp/"を入力

コメント